
Croissance microbienne - Suivi de croissance (M)
Objectif : realiser un suivi de croissance de différents micro-organismes dans les mêmes conditions opératoires :
- température de 37 °C en bain thermostaté
- aérobiose par agitation manuelle
- bouillon LB (composition pour 1 L)
- 10 g de tryptone = peptone trypsique : peptides et acides aminés issus de l'hydrolyse de protéines par la trypsine, source d'azote, carbone et énergie
- 5 g d'extraits de levures : source de facteurs de croissance (vitamines, bases azotés, acides aminés) et sels minéraux
- 10 g de chlorure de sodium : maintien de la pression osmotique du milieu
- pH final de 7,5
- souches tests :
- Escherichia coli, bacille polymorphe à Gram négatif, oxydase négatif, de la famille des Enterobacteriaceae
- Staphylococcus epidermidis, coque à Gram positif, catalase positive
- Saccharomyces cerevisiae, mycète de levure, eucaryote unicellulaire
La croissance du micro-organisme sera réalisée dans une fiole d’Erlenmeyer de 250 mL contenant 95 mL de bouillon LB, à 37 °C dans un bain thermostaté, sous agitation douce manuelle.
Matériel biologique
-1 tube de préculture en bouillon LB après 14 heures à 37 °C
Autre matériel
-1 fiole d’Erlenmeyer contenant 95 mL de LB stérile (préchauffé à 37 °C)
-3 à 4 pipettes graduées stériles de 1 mL
-16 semi-microcuves de contenance 1 mL et papier Parafilm®
-1 tube de LB stérile 10 mL
-2 géloses nutritivesordinaires
Avant de commencer la croissance
-À partir du bouillon LB stérile de l’Erlen : prélever stérilement 1 mL et l’introduire dans une microcuve pour l’ajustage au zéro du turbidimètre. Parafilmer.
-Conserver ce blanc réactif pour toute la durée de la manipulation.
-À partir de la préculture fournie : vérifier immédiatement la pureté de cette suspension par un isolement sur gélose nutritive ordinaire (GNO) ou TSA (Sabouraud + chloramphénicol si levures).
Suivi de croissance
-Vérifier la température du bain thermostaté.
-Ajouter stérilement 5 mL de préculture dans le bouillon LB préchauffé
-Reboucher et agiter délicatement. Déclencher le chronomètre.
-Immédiatement : prélèvement à t0 = 0 min :
- Prélever une fraction aliquote de la culture à l'aide d'une pipette stérile de 1 mL, et l’introduire stérilement dans une microcuve.
- Laisser la pipette dans l'Erlenmeyer, coincée avec le bouchon de coton.
- Mettre l'Erlenmeyer au bain thermostaté à 37 °C et agiter très régulièrement pour favoriser l'aérobiose.
- Mesurer l’atténuance D600 à t0 min contre le blanc réactif.
- Suivi cinétique de croissance pendant 120 à 180 minutes :
- Toutes les 10 minutes exactement, prélever stérilement 1 mL de culture.
Bien laisser la pipette coincée dans l’Erlenmeyer après ce prélèvement. - Mesurer immédiatement l’atténuance contre le blanc réactif.
- Lorsque l'atténuance mesurée devient supérieure à l'atténuance limite (cf. phase I et son exploitation), diluer directement en micro-cuve.
Noter le facteur de dilution et calculer l’atténuance corrigée. - Vérifier la pureté de la culture en fin de croissance : réaliser un isolement sur GNO ou TSA.
- Toutes les 10 minutes exactement, prélever stérilement 1 mL de culture.
Chronogramme de la procédure opératoire :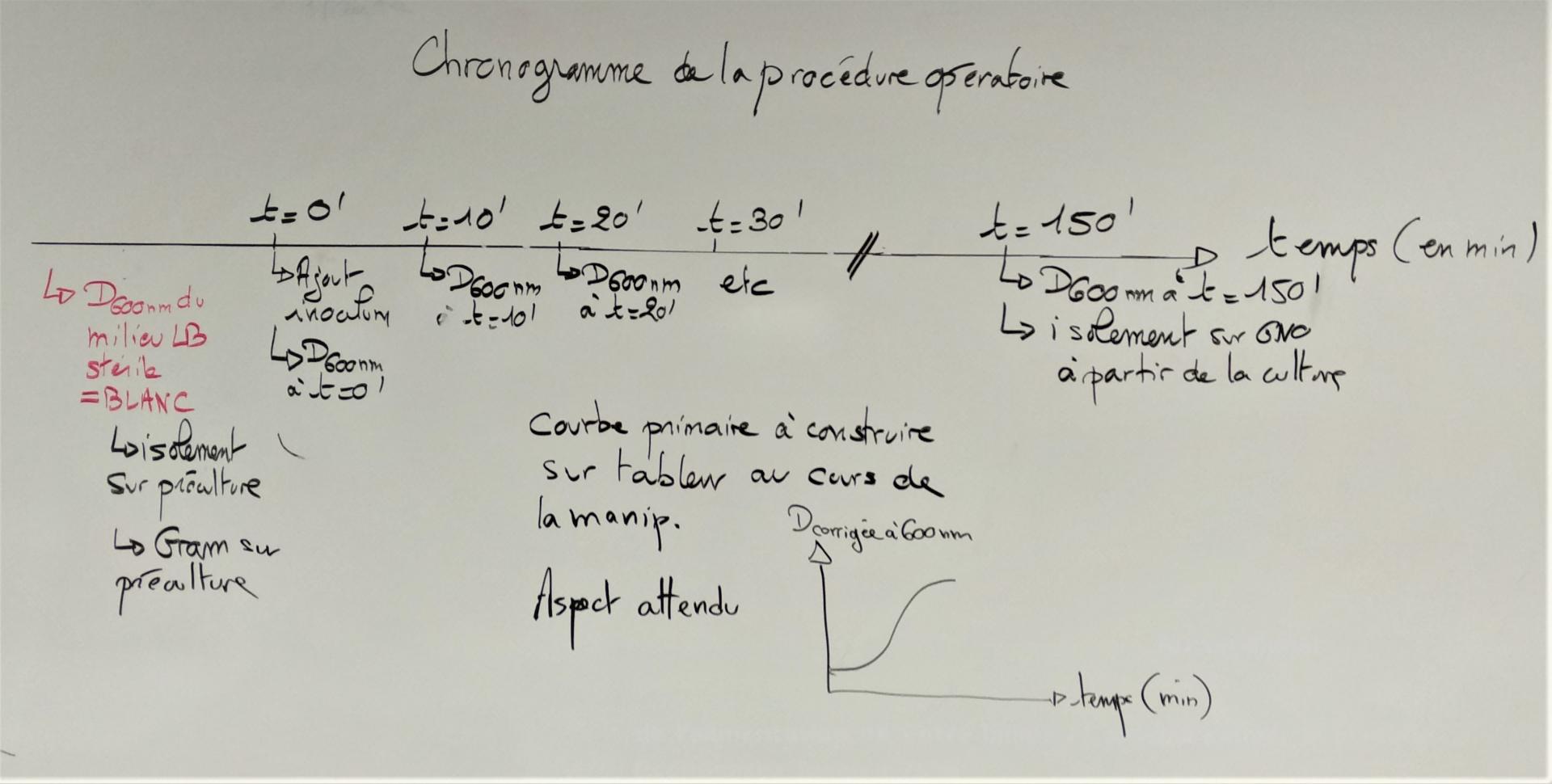
Le suvi de croissance permet de déterminer :
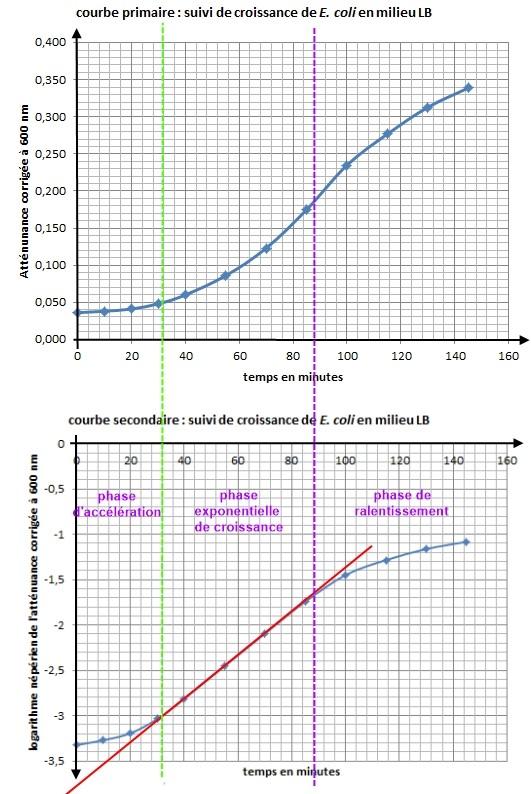
- les phases de la croissance (phase de latence, d'accélération, exponentielle, de ralentissement, stationnaire, de déclin)
- les paramètres cinétiques de la croissance : vitesse spécifique de croissance en phase exponentielle µexpo ou µmax, temps de génération G :
- la vitesse spécifique maximale de croissance µmax (ou µexpo ou QX) est déterminée pendant la phase exponentielle, phase pendant laquelle il y a une relation de proportionnalité entre l’augmentation de la biomasse et le temps sur la courbe secondaire ln (X) = f(t) (X représentant la biomasse, évaluée par la mesure de l'atténuance à 650 nm : D650)
µmax est la quantité de cellules produites par unité de temps, ramené au nombre de cellules.
Sur la courbe secondaire ln X = f(t) : µmax = (ln D2 - lnD1) / (t2 - t1) qui est le coefficient directeur, calculé en choisissant des points les plus éloignés possibles, pris sur la tangente à la phase exponentielle de croissance.Signification de µmax : admettons que = 1,38 h-1
cela signifie que, en 1 h, le rapport (nombre de cellules produites / nombres de cellules) dans l’erlen est de 1,38, ou encore qu’on a 138 cellules produites pour 100 cellules au départ. - le temps de génération G est l’intervalle de temps permettant le doublement de la population ou encore le temps que met une cellule pour se diviser.
Il s’exprime en h ou en min et peut être déterminé de trois façons différentes :
- par le calcul : considérons que (t2 - t1) correspond à G ; dans ce cas, la biomasse ou l'atténuance aura doublé lorsque le temps de génération se sera écoulé :
D2 = 2D1 soit, en conversion logarithmique : ln D2 = ln2D1
Dans l'équation du calcul du pente, cela donne : µmax = (ln2D1 - lnD1) / G soit µmax = (ln2 + lnD1 - lnD1) / G qui se simplifie donc en µmax = ln2 / G
Il ne reste qu'à isoler le temps de génération : G =ln2 / µexpo - par détermination graphique sur la courbe secondaire: si on part d'un temps t1 et qu'on relève la valeur correspondante de ln D1, la population aura doublé après une durée égale à G ; on aura D2 = 2 D1 donc ln D2 = ln (2 D1) = ln 2 + ln D1 = 0,7 + ln D1 .
Pour ln X2 ainsi calculé on relève t 2, on peut calculer G = t2 – t1 - par détermination graphique sur la courbe primaire : si on part d'un temps t1 et qu'on relève la valeur correspondante de D1, la population aura doublé après une durée égale à G ; on aura D2 = 2 D1 ; il suffit de reporter sur la courbe et déterminer le temps t2, puis on peut calculer G = t2 – t1
ATTENTION : ces paramètres ne peuvent être calculés qu'en phase exponentielle de croissance
Ces deux paramètres sont propres à une souche microbienne donnée et pour une même souche, ils peuvent varier en fonction des conditions de cultures (composition du milieu, pH, température, pO2 …)
Erlenmeyer contenant 95 mL de bouillon LB stérile inoculé avec 5 mL de préculture d'E. coli

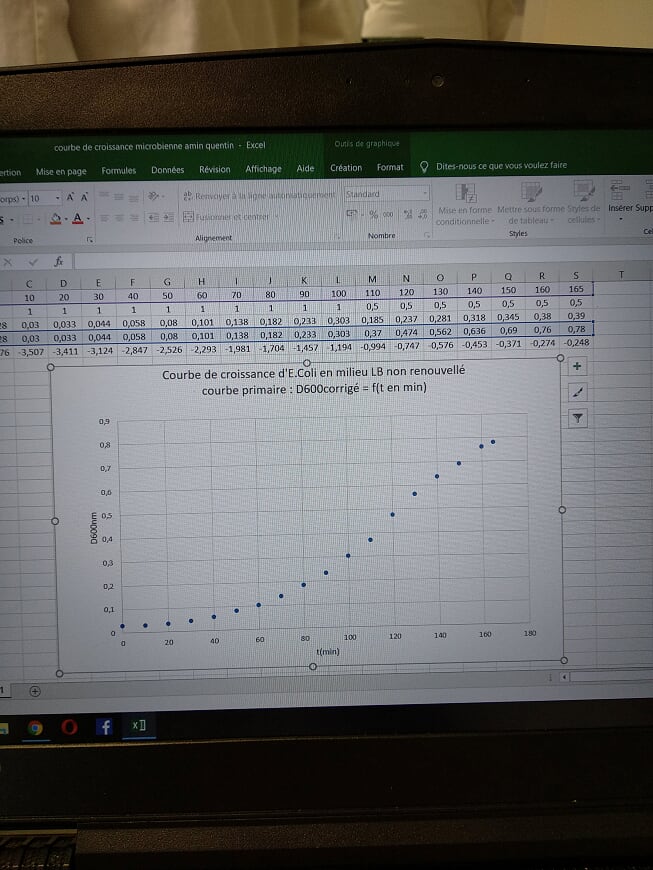
Construction en temps réel du nuage de points représenant l'atténuance corrigée à 600 nm (contre milieu LB stérile) en fonction du temps de croissance en minutes.
Courbes obtenues par un autre binôme :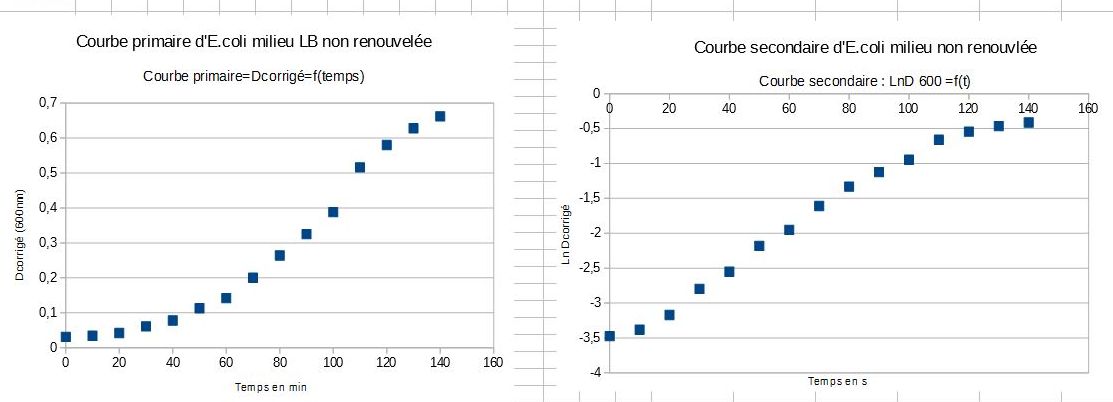
Exploitation graphique de la courbe primaire et de la courbe secondaire
Contrôle par observation microscopique (frottis fixé coloré au Gram) des caractères morphologiques la souche mise en oeuvre dans le suivi de croissance (Escherichia coli)
Notons une coloration bipolaire (les extrémités du bacilles apparaissent davantages colorées en rose que le centre, pouvant mener à une interprétation erronée de diplocoque) sur ce frottis fixié coloré au Gram, montrantdes formes coccobacilles et bacilles allongés, à bouts ronds d'environ 1,5 µm de long.

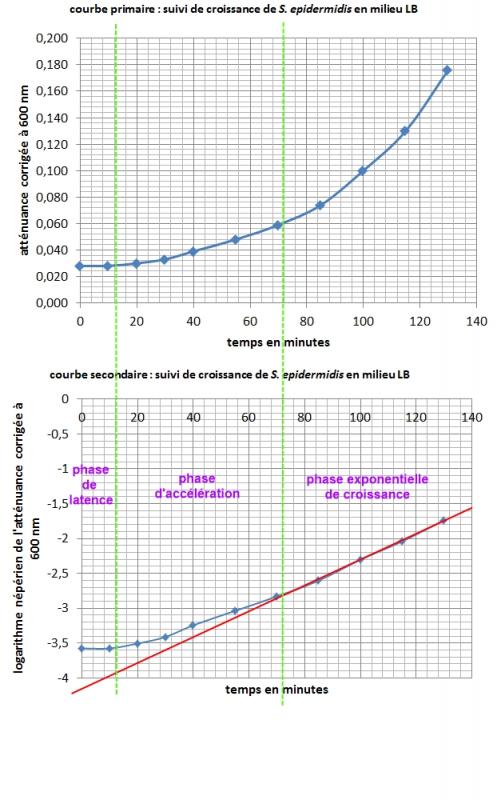
Calculer la vitesse spécifique maximale de croissance µmax (ou µexpo ou QX) en phase exponentielle, p
Calculer le temps de génération G.
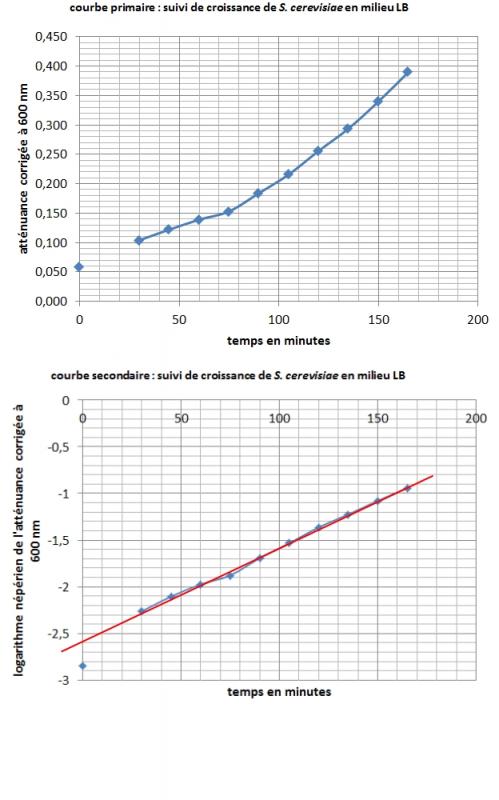
Calculer la vitesse spécifique maximale de croissance µmax (ou µexpo ou QX) en phase exponentielle, p
Calculer le temps de génération G.
Les phases de la croissance microbienne
|
PHASE |
DESCRIPTION |
|
Phase de latence |
Phase facultative : correspond à une phase d'adaptation des micro-organismes au milieu (temps pour synthétiser les enzymes catalysant la dégradation d substrats nouveaux du milieu, réparations cellulaire, synthèse d’ATP, de cofacteur). |
|
Phase d'accélération |
Phase facultative : correspond au démarrage de la croissance ; les micro-organismes se multiplient de plus en plus activement. Le temps de génération G atteint progressivement sa valeur minimale. |
|
Phase exponentielle de croissance |
Les micro-organismes se multiplient à leur vitesse spécifique µ la plus élevée Rem : c’est pendant cette phase qu’il y a production d'antibiotiques et de toxines. |
|
Phase de ralentissement |
Les bactéries continuent à se multiplier mais de moins en moins vite ; la vitesse spécifique de croissance diminue progressivement, ce qui peut s’expliquer par :
|
|
Phase stationnaire |
La biomasse est stable, soit parce que :
|
|
Phase de déclin |
Certaines bactéries meurent et sont lysées. Leur nombre diminue. Dans certains cas des bactéries survivantes peuvent se multiplier au dépend des substances libérées par la lyse bactérienne. |
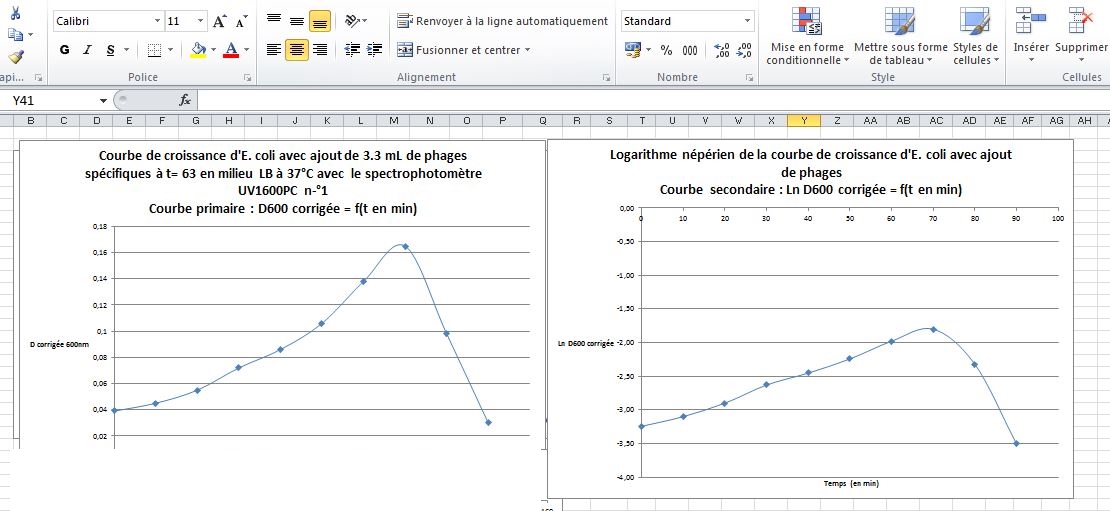
Bonus de manipulation : deux binômes ont inoculé dans l'Erlenmeyer, au cours de la phase exponentielle de croissance, 2 mL d'ue suspenbsion phagique conservée depuis une activité précédente.
Cette suspension de phages a été obtenue à partir d'un prélèvement d'eau de rivière en aval d'une station d'épuraton, orélèvement dont on espérait mettre en évidence et dénombrer des phages spécifiques de diverses bactéries.
Un type de phage infectant spécificiquement E. coli avait été obtenu ; les phages avaient alors subi une étape d'amplification in vivo et, après filtration, une suspension phagique concentrée avait été obtenue et conservée.
Construction de la courbe de croissance en temps réel ; la suspension phagique a été noculée à t = 60 minutes.
Ci-desosus comparaison à gauche, l'Erlen contenant le bouillon LB après 2h30 de croissance d'E. coli et à droite, le même bouillon après 1h30 de croissance d'.E coli suivi de 1h10 d'infection phagique !
Date de dernière mise à jour : 14/02/2024
Ajouter un commentaire