
Restriction enzymatique de l'ADN du phage lambda
Un marqueur de taille ADN (« DNA marker » en anglais) est un mélange de molécules d’ADN linéaires, de tailles connues et variées qui sont bien séparables sur un gel d’agarose par électrophorèse.
Un marqueur de taille est utilisé au cours de chaque électrophorèse en gel d’agarose de solutions d’ADN à analyser. Il permet, par comparaison entre la distance de migration des différentes bandes d’ADN du marqueur de taille et la distance de migration d’une bande d’ADN de taille inconnue, d’estimer la taille de cette bande d’ADN inconnu.
Un des marqueurs de taille commercial utilisé au laboratoire : Gene ruler 1 kb DNA ladder (Fischer Scientific) :

Combien de fragments d'ADN linéaire contient ce marqueur de taille ?
Quel est le pourcentage d'agarose du gel ayant permis la séparation électrophorétique de ces fragments ?
Ce marqueur de taille est-il adapté pour séparer des fragments d'ADN linéaires
- dont la taille serait comprise entre 10 000 et 50 000 pb (pb = paires de bases) ?
- dont la taille serait comprise entre 50 et 250 pb ?
Est-ce que certains fragments sont plus facilement repérables sur le gel après révélation ? Si oui, lesquels ?
Deux types de marqueurs de taille peuvent être obtenus :
-obtenus par mélange d'ADN d’origines différentes ;
-obtenus par restriction enzymatique (syn. : digestion, coupure) enzymatique d’un ADN (phagique ou plasmidique) à l'aide d'enzymes de restriction (endonucléases)
Objectifs :
- Prévoir par bio-informatique la taille des fragments attendus pour une restriction enzymatique d'un ADN de taille connu par deux enzymes de restriction différentes (restriction virtuelle dite restriction in silico).
- Réaliser une restriction enzymatique simple afin de découvrir les caractéristiques de enzymes de restriction.
- Vérifier si il est possible de fabriquer notre propre marqueur de taille par restriction d’un ADN phagique (ADN du bactériophage lambda).
- Vérifier par électrophorèse sur gel d’agarose si les fragments obtenus correspondent à la prévision théorique.
Restriction enzymatique de l'ADN du bactériophage lambda
Compléter le tableau de manipulation suivant (tous les calculs doivent être justifiées par les équations aux grandeurs, aux unités, aux valeurs numériques) en respectant la chonologie suivante :
- Rechercher le volume final contenu dans le microtube.
- Calculer Vsol ADN : volume de solution d’ADN du phage lambda à mettre dans chaque microtube.
- Calculer le volume de tampon 10X (i.e. concentré 10 fois) HindIII et AvaI (= Eco88I) à mettre dans chaque microtube, sachant qu'il doit être ramené à 1X (concentré 1 fois)
- Compléter le volume d’eau à ajouter (qsp) dans chaque microtube.
- Valider que le volume d’enzyme à 10 U · µL-1 ajouté dans les microtubes (1 µL) permet d’apporter 10 U pour 1 µg d’ADN du phage lambda
Donnée : il est recommandé d’avoir entre 10 et 20 U par µg d’ADN avec un volume d’enzyme au maximum égal à 1/10 du volume total.
|
Microtubes |
0 |
H |
A |
|
Tampon 10X pour Hind III (µL) |
0 |
|
0 |
|
Tampon 10X pour Ava I (µL) |
0 |
0 |
|
|
Eau qualité BM qsp 20 µL (µL) |
|
|
|
|
Solution d’ADN l à 250 ng · µL-1 apportant 500 ng (µL) |
|
|
|
|
Enzyme Hind III Fastdigest à 10 U · µL-1 (µL) |
0 |
1 |
0 |
|
Enzyme Ava I (=Eco88I) Fastdigest à 10 U · µL-1 (µL) |
0 |
0 |
1 |
|
Durée d’incubation à 37 °C (min) |
5 |
5 |
5 |
|
Durée d’incubation à 65 °C (min) |
15 |
15 |
15 |
En utilisant le document suivant, justifier le tampon à utiliser :
- Pour réaliser une simple restriction de l’ADN l par l’enzyme de restriction HindIII.
- Pour réaliser une simple restriction de l’ADN l par l’enzyme de restriction AvaI (=Eco88I)
- Pour réaliser une double restriction de l’ADN l par HindIII et AvaI.
Activité des enzymes de restriction HindIII, EcoRI et AvaI (Eco88I) dans 4 tampons
|
Enzymes de restriction |
% d’activité de l’enzyme dans les tampons 1, 2, 3 et 4 |
|||
|
1 |
2 |
3 |
4 |
|
|
HindIII |
25 |
75 |
100 |
50 |
|
EcoRI |
25 |
100 |
50 |
50 |
|
AvaI |
10 |
100 |
25 |
100 |
Note : FastDigest enzymes are an advanced line of restriction enzymes for rapid DNA digestion. All FastDigest™ enzymes are 100 % active in the universal FastDigest and FastDigest Green buffers and are able to digest DNA in 5-15 minutes. 1 µL of FastDigest is formulated to digest up to 1 µg of lambda DNA in 5 min and 1 µg of plasmid DNA in 15 min
Les enzymes utilisées pour cette restriction (référence FastDigest) nécessitent-elles un des tampons 1 à 4 ? (Traduire la note pour répondre à la question)
Préciser le rôle du microtube 0. Prévoir son résultat.
Indiquer le rôle de l’incubation à 65 °C.
Calculer le volume de solution (bleu) de dépôt 5X à rajouter dans les échantillons sachant que dans chaque puit du gel seront déposés 25 µL de solution contenant le mélange réactionnel de restriction + la solution de dépôt.
A l’aide du document suivant :
- représenter les extrémités des fragments de restriction obtenus après digestion par les enzymes de restrictions AvaI (Eco88I) et HindIII
- indiquer la nature de ces extrémités : franches, cohésives -ou collantes- 5'- ou 3'-sortantes.

Aide : repérer les flèches qui indiquent les points de coupure, "découper" la séquence palindromique en séparant les bases azotés, et ne pas oublier de rajouter les extrémités 5' et 3' libérées après la coupure.
Les séquences reconnues par les enzymes de restriction (appelées sites de restriction) sont de courtes séquences (6 à 10 nuécléotides) palindromiques. Emettre une hypothèse sur la stucture des enzymes de restriction, leur permettant de couper simultanément au même endroit sur chaque brin d'ADN.
Simulation par bio-informatique de la restriction enzymatique
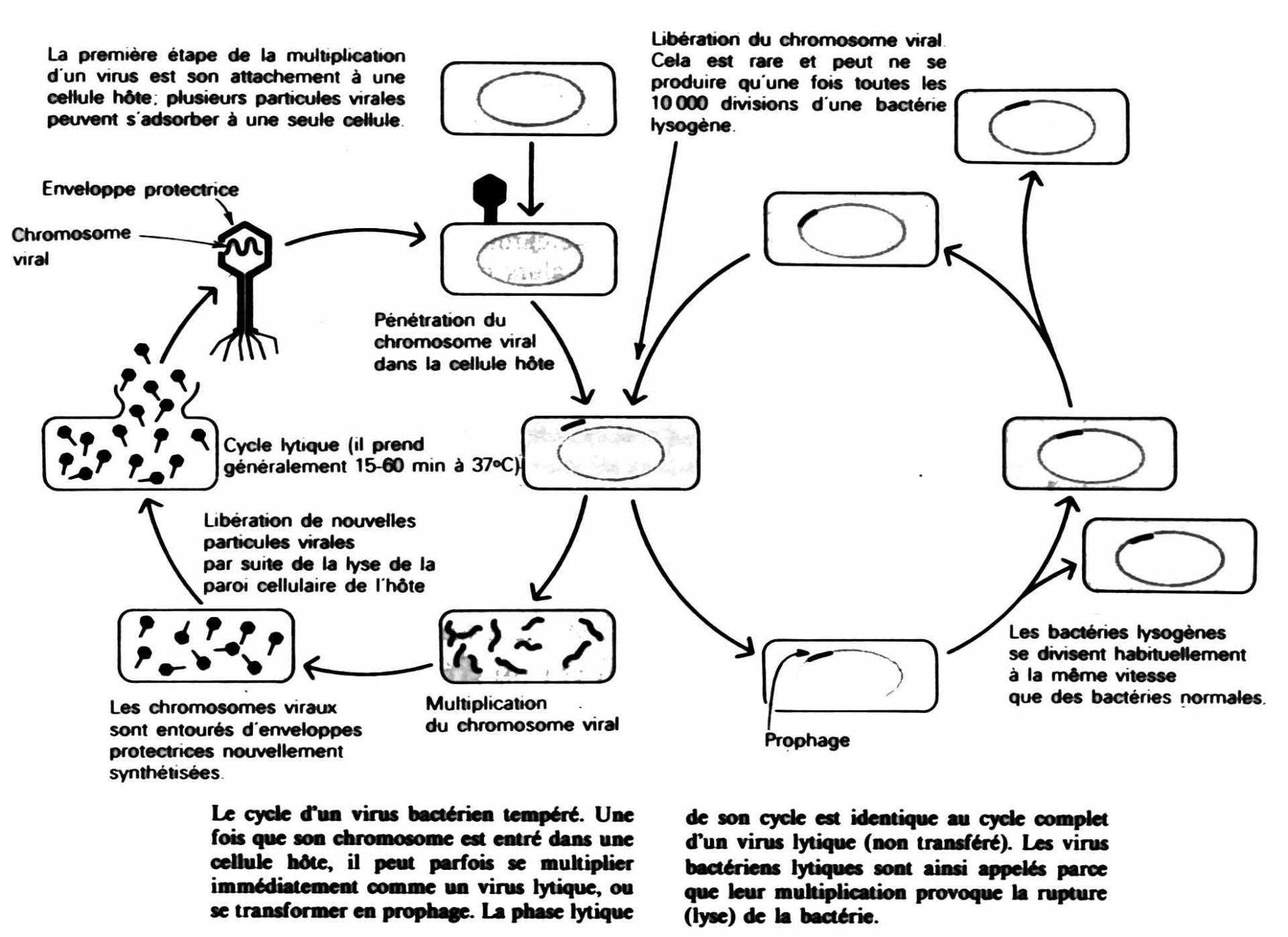
L'ADN utilisé comme substrat de la digestion enzymatique est celui du bactériophage lambda : son génome est constitué d'ADN double brin de 48 502 pb (soit 48,5 kilobases, kb). Dans la particule virale, l’ADN est linéaire, mais ce dernier possède des extrémités cohésives (12 nucléotides simple brin complémentaires).
Au cours de l’infection, le phage lambda va injecter son génome dans le cytoplasme de la bactérie. Les extrémités cohésives peuvent alors s’apparier pour former un génome circulaire et ainsi permettre sa réplication.
Schéma du cycle de réplication du bactériophage lambda : ce virus bactérien peut adopter un cycle lytique ou un cycle lysogénique (il peut rester à l'état latent à l'intérieur d'une bactérie, son ADN viral étant intégré à l'ADN bactérien). C'est un phage dit tempéré
Utiliser le site Nebcutter https://nc3.neb.com/NEBcutter/ pour réaliser virtuellement :
- la restriction enzymatique de l'ADN du phage l par Hind III
- la restriction enzymatique de l'ADN du phage l par Ava I (Eco88I)
- la double restriction zymatique de l'ADN du phage l par Hind III et Ava I (Eco88I)
Exemple de résultat de simulation obtenu :







Peut-on supposer que la séparation électrophorétique des fragments d'ADN issus de la restriction de l'ADN du phage lambda par l'enzyme HindII permettra de bien séparer tous les fragmens en bandes bien distinctes ? Si non, expliquer pourquoi
Préparation du gel d'agarose et de l'électrophorèse
Calculer le volume de tampon TBE 10X à utiliser pour préparer 1 L de tampon TBE 1X.
Calculer la masse de poudre d’agarose à peser pour préparer un gel d’agarose à 0,8 % (m/V) occupant une surface de 7 x 12 cm sur une hauteur de 0,5 cm (dimensions du support de coulage de gel)
Préparation du gel :






Préparation des supports de coulage :

Coulage du gel :

Séparation électrophorétique sur gel d'agarose
Dépot des échantillons dans les puits du gel d'agarose : astuces pour réaliser un dépôt correct :
- disposer un fond noir sous le gel afin de bien visualiser les puits,
- poser les deux coudes sur la paillasse de part et d'autre du gel,
- se positionner bien au-dessus du gel pour voir les puits,
- diriger l'extrémité de la pipette à piston à l'aide d'un ou deux doigts de la main faible (celle qui ne tient pas la pipette)
- pour vérifier si le cône de la pipette est bien dans le puits, un très léger mouvement avant/arriere peut être réalisé : le gel doit se déplacer délicatement
- attention : les gels sont fragiles, surtout si ils sont à seulement 0,7 % d'agarose
- pousser le piston de la pipette d'un mouvement lent, uniforme, sans jamais le relacher sans quoi l'échantillon va être ré-aspiré dans le cône

Migration électrophorétique (90 V, 500 mA, 75 min) :
Arrêt de la migration :
On notera un problème de dosage avec le BBT qui sert de marqueur de front de migration : quantité trop importante dans la solution de loading, qui risque de masquer les fragments de restriction.
Révélation des fragments d'ADN et scan du gel
Deux types d'agents révélateurs peuvent être utilisés :
- des colorants spécifiques de l'ADN (non toxiques, non cancérigènes) mais dont la qualité de révélation est moindre
- des agetns intercalants qui se fixent entre les bases azotés (toxiques, cancérigènes) mais qui offrent une meilleure qualité de révélation
Exemples :
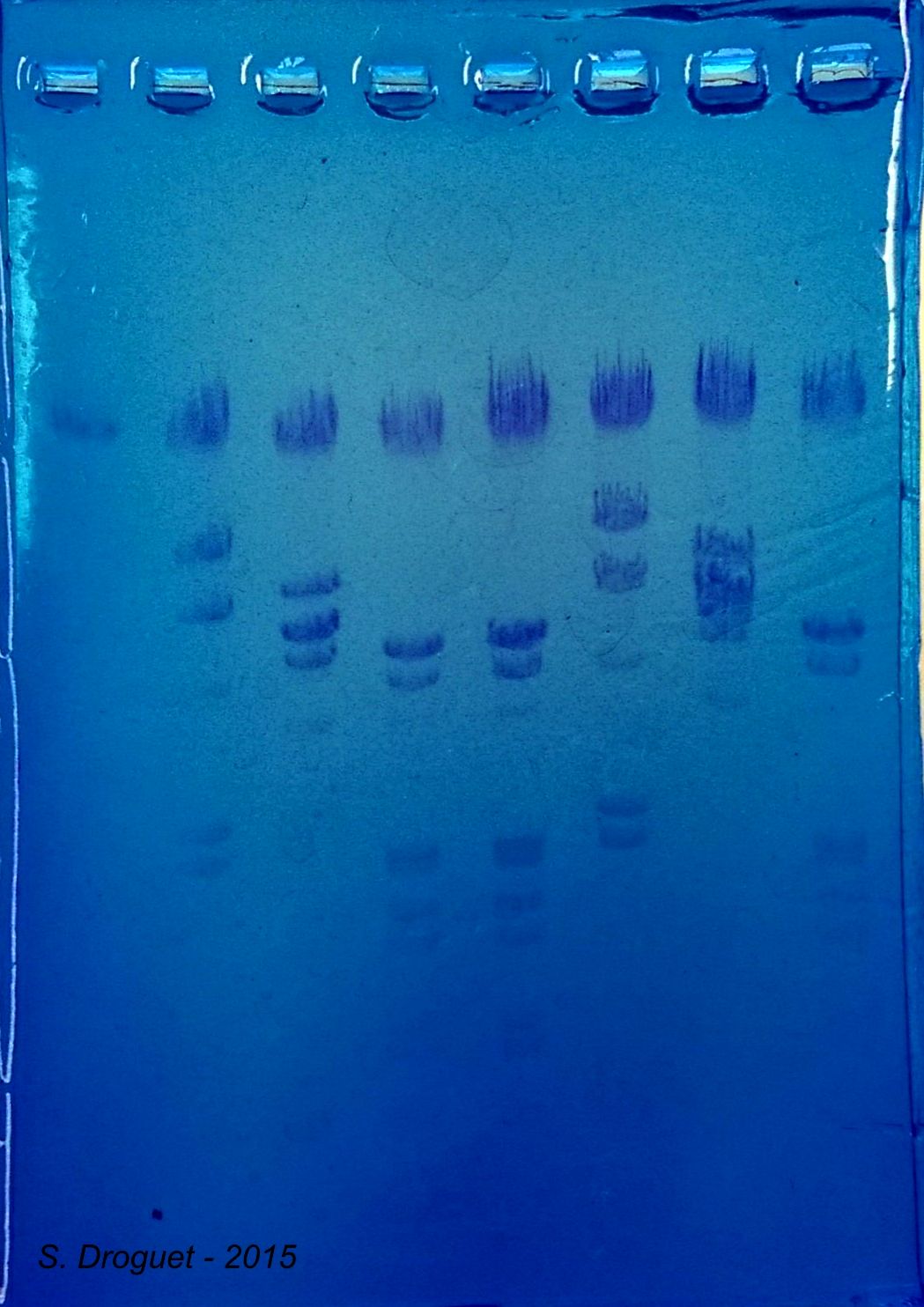
Révélation au bleu azur (photo réalisée en 2015, avec un smartphone, donc qualité non optimale) :
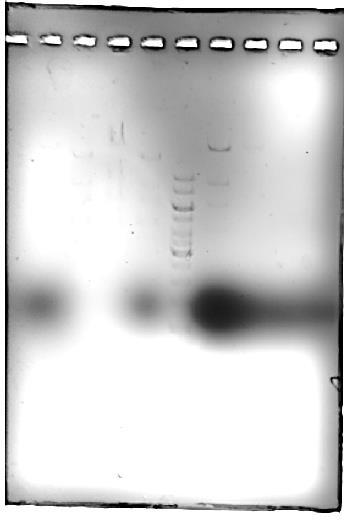
Révélation avec un agent intercalant (Gelred) (photo réalisée avec le lecteur de gel Biorad GelDoc EZ, donc qualité optimale, sans post-traitement) :
Le colorant utilisé cette année porte le nom de Fast Blast DNA Stain ; extraits de la notice technique :






Les gels sont scannés avec un lecteur de gel Biorad GelDoc EZ

Lecture du gel après scan avec le GelDoc EZ ; la décoloration n'est pas optimale (bruit de fond important) et le marqueur de front de migration masque certains fragments
:
Lecture de la piste "Marqueur de taille"
- Combien de fragments sont visibles ?
- Combien devrait-on en voir ?
- Légender sur le coté la taille des fragments du marqueur de taille (Gene Ruler 1 kb DNA ladder)
- Pourrait-on envisager de tracer une droite d'étalonnage à partir de cette piste, qui permettrait de déterminer la longueur de fragments inconnus par comparaison avec cette droite ?
Eléments d'explications :
- un problème de concentration de la solution de charge (utilisée comme étant à 5X mais en réalité à 6X, et avec une possibilité de concentration excessive de BBT) masque les petits fragments
- le révélateur utilisé l'était pour la 1ère fois ; son utilisation reste à optimiser
- Pour tracer la droite d'étalonnage, il faut mesurer la distance de migration de chaque fragment par rapport au point de départ (puits). Connaissant la taille de chaque fragment, on pourrait construire un graphe : taille du fragment = f(distance de migration), qui permettrait de se rendre compte que la relation n'est pas linéaire. Une fonction mathématique, déjà utilisée lors de l'étude de la croissance microbienne, permet alors de linéariser la courbe précédente, en calculant le logarithme décimal de la taille de chaque fragment.
On trace alors la droite log (taille des fragments en pb) = f(distance de migration)
Lecture de la piste "H" (à droite du marqueur de taille) :
- Combien de fragments sont visibles ?
- Combien devrait-on en voir d'après la restriction in silico ?
- La restriction a-t-elle été totale ou partielle ?
- Pourquoi ne voit-on pas tous les fragments ?
- Estimer la taille des fragments observables.
Eléments d'explications :
- dans le cas d'une digestion partielle, on observerait de nombreux plus nombreux fragments que ne l'indique la simulation informatique ; en effet, certaines molécules d'ADN ne pourraient subir que 2 coupures au lieu de 6
- la quantité d'ADN du phage réellement déposée est inférieure à celle indiquée dans le tableau de préparation ; la solution d'ADN de phage était à 150 ng.mL-1 au lieu de 250, donc la masse totale d'ADN déposée n'était que de 300 ng au lieu de 500 ng
- le révélateur se fixe spécifiquement sur l'ADN ; aussi, les petits fragments fixent moins de révélateur, et sont donc moins visibles.
Exemple d'annotation de l'élétrophorégramme :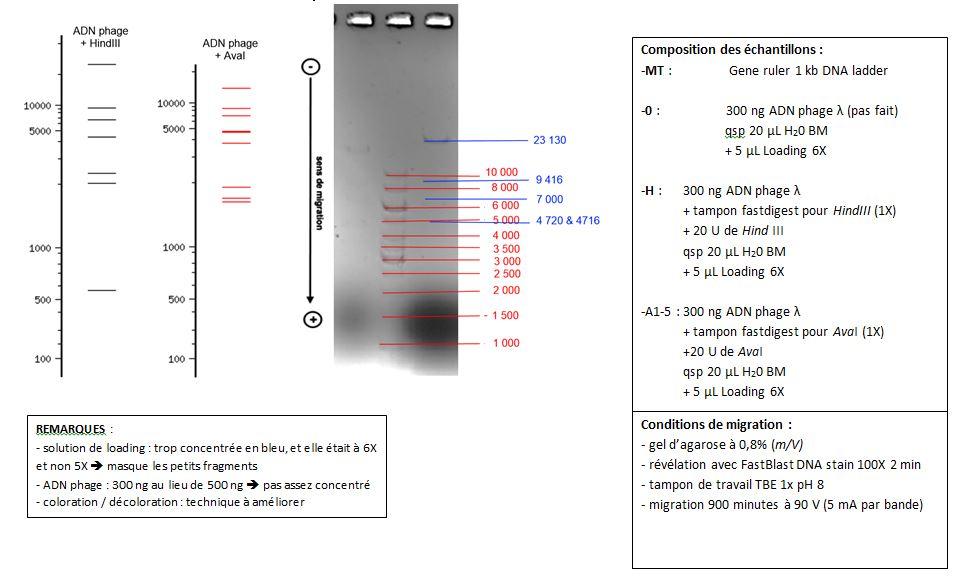
Manipulation refaite la semaine prochaine... à suivre
Résultat brut de la seconde réalisation
Le témoin négatif de restriction a été réalisé (puis "0", contenant uniquement l'ADN du bactériophage lambda) :



Date de dernière mise à jour : 02/06/2024
Ajouter un commentaire